生物の周期的な連続性は、生殖の現象における世代間のつながりを見つけます。
生殖は、進化のスケールのさまざまなレベルで、野菜と動物の王国のさまざまな影響で、さまざまな生物種で、単独で論文全体を正当化するようなさまざまなメカニズムで実行されます。
生殖現象の最初の分類は、単細胞生物と多細胞生物を区別する必要があります。これは、前者でのみ細胞分裂が生殖と一致するためです。
多細胞では、生殖はアガミックまたはセクシュアル(またはガミック)である可能性があります。
比較的頻度の低いアガミック生殖は、有糸分裂のメカニズムに基づいているため、種の変動性は、突然変異の繰り返し発生にかなり委ねられています。
横分体形成、胞子形成などのさまざまなメカニズムもありますが、植物では、農業でよく知られている再生形態(挿し木、層状化など)が見られます。
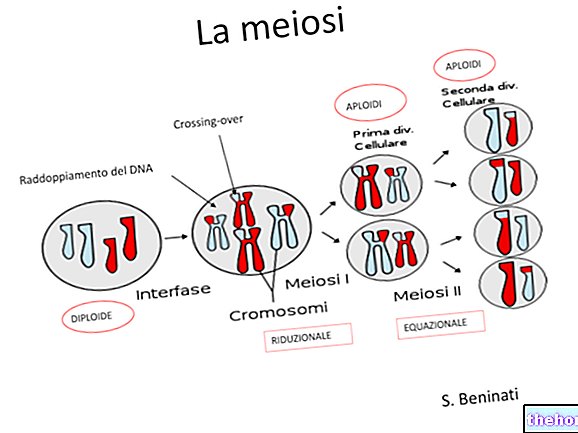
しかし、より高い形態で最も普及している生殖メカニズムは、減数分裂の出現、配偶子の形成、および接合子におけるそれらの融合(受精)に対応する性的メカニズムです。
原始種では、配偶子は形態学的に区別されていません。この場合、私たちは等配偶子について話します。ただし、記号(+)と(-)で識別される2つの一連の配偶子が常に存在し、受精は「反対の符号の配偶子間の遭遇からのみ発生します。したがって、生物学的な違いがあり、まだ形態学的に表現されていません。
進化のスケールが進むにつれて、形態学的および機能的な分化が現れます。そこでは、一般に豊富な予備物質(代謝的に独立するまで胚の発達を確実にする重膜または子牛)とタイプの雌の配偶子があります。男性の配偶子は女性の配偶子に到達するための運動性に恵まれています。配偶子は常に半数体であり、減数分裂の結果です。それらの融合は二倍体の接合子を生じさせます。
減数分裂と接合子の間では、一連の一倍体細胞世代が経過する可能性があります。ちょうど、接合子と減数分裂の間では、世代交代によって表されるさまざまな種類の生成サイクルで、一連の二倍体細胞世代が経過する可能性があります。
(二倍体生物を含む)二倍体種は、配偶子減数分裂を特徴とします。減数分裂は、直接配偶子を生成し、それが融合することにより、二倍体状態を即座に再構成します。これは、人間を含む後生動物の一般的なケースです。
配偶子形成
人間で起こる減数分裂の生殖を考慮して、減数分裂が配偶子形成(配偶子の形成)にどのように適合するかを明らかにしてみましょう。
男性と女性の配偶子形成(精子形成と卵形成と呼ばれる)の胚発生では、体を形成する運命にある細胞(体細胞系列)と配偶子を生成する運命にある細胞(生殖細胞系列)の間に初期の分化があります。生殖系列の最初の細胞はプロトゴンと呼ばれます。男性または女性の意味での性腺の分化に伴い、生殖細胞はそれぞれ精原細胞と卵祖細胞に分化します。
精子形成を見ると、精原細胞には一連の細胞世代があり、それは生涯続くことがわかります。このようにして継続的に産生される精原細胞の一部のみが通常の有糸分裂周期と異なり、代わりに減数分裂が始まります。
減数分裂が始まる生殖細胞(複製、次に最初の分裂)は一次精母細胞と呼ばれます。その分裂は2つの二次精母細胞を生じさせ、それは2番目の分裂で合計4つの精母細胞を生じさせます。
染色体キットの減少を、4nの一次精母細胞(複製後、ホモログの各ペアに4つの染色体があります)から2nの二次精母細胞およびnの精母細胞に段階的に変化させることができます。したがって、精母細胞はすでに半数体ですが、まだ成熟した配偶子ではありません。半数体型細胞の構造から、機能的成熟(精子組織形成と呼ばれる)は精母細胞を精母細胞、つまり成熟した男性の配偶子に変換します。
女性の配偶子形成(または卵形成)にはいくつかの違いがあります。まず第一に、準備される配偶子の数ははるかに少ないです。約5X 105のオボゴンが、ヒト種の女性の性腺で作られていると推定されています。これらのうち、約35年間の出産期間の間、通常は月に1つの卵胞にのみ影響を与えるサイクルで、約400のみが卵胞の成熟とその後の落胆に関心があります。
両性で準備された配偶子の数の違いは、すでに述べた機能と行動の違いに対応しています。精子は小さく、可動性があり、卵子を探す必要性とそれを見つける可能性が低いことに関連して多数あります。胚に予備物質を保証する機能と、体内受精によってそれらに提供される保護に関連して、大きく、不活性で、少数です(当然、特に体外受精では、卵子もより多くなければなりません)。
配偶子に予備物質を提供する必要性は、「卵形成において、減数分裂の停止段階の存在に対応し、その間に染色体は部分的に脱スパイラル化されます。次に、いわゆる「羽毛染色体」を観察します。外向性の数は、配偶子の合成に関与する遺伝子が抑制解除される特性を識別します。
女性の配偶子に必要な数が少ないことは、減数分裂によって生成された4つの一倍体細胞のうち、1つだけが予備物質全体を受け取って配偶子になり、他の3つ(染色体または極体)は染色体物質は、接合子や胚を生じさせることができず、退行する運命にあります。
受精
受精、つまりオスとメスの配偶子の出会いは、非常に異なる方法で行うことができます。動物界では、体外受精(環境リスクにさらされているため、雌雄で必然的に非常に多い配偶子)から体内受精への移行が観察されます。これは、親の世話が母親と胎児の間の哺乳類の代謝関係にさらに関連しています。 。
受精は、異性の配偶子同士の出会いが起こったら、特異性と独自性という2つの条件を保証して行われなければなりません。つまり、精子が卵子と同じ種であり、最初の精子が入ると、他の精子が入らないようにする必要があります。
特異性は、先体と卵子の表面の生化学的特性によって保証されます。実際、«フェルチリシン»と«アンチフェルチリシン»の間の反応の話があり、その特異性は酵素と基質の間の遭遇のそれに匹敵します。
受精の独自性は、最初の特定のフェルチリシン/抗フェルチリシン反応の瞬間に始まる卵子の表面構造の変更(「皮質反応」)によって保証されます。この反応の後、卵子の膜が変化し、それに到達した他の精子は特定の受精反応を開始できなくなります。
受精後、精子の尾は卵子の外側に残り、染色体物質が卵子に浸透します。これは「雄の前核」と呼ばれ、卵子の「雌の前核」と結合して接合子の二倍体核を形成します。