ただし、「説明のない同様のステートメントは、特にトピックを理解するために必要な知識を持っていないすべての人にとって、誤解を招く可能性がある」ことは論理的です。
極端なものから別の極端なものまで、この「新しいマントラ」を誤解したり、さらに別の代替栄養システムを促進するための言い訳として使用したりする人はたくさんいます。
この記事では、脂肪が炭水化物の火で燃えることの意味を理解しようとしますが、とりわけ、この概念が痩身プロセスにどのように影響するかを理解しようとします。
ただし、「正しい」と認識される唯一の痩身方法は、バランスの取れた方法、つまり、ケースのすべてのトラップを尊重する方法であると予想されます。同じカロリーで、エネルギー主要栄養素のすべての分解(炭水化物、タンパク質、脂肪)はほぼ同じ結果をもたらします。重要なのはカロリーバランスであり、これは明らかに負である必要があります。
ただし、適用性や運動能力、特に持久力スポーツのパフォーマンス、除脂肪体重の維持、筋力やボディービルなどには大きな違いがあります。
しかし、脂肪が炭水化物の火で燃えるとはどういう意味ですか?簡単に言えば、生化学的に言えば、脂肪酸の細胞酸化はブドウ糖なしでは成し遂げられませんが、以下で可能な限り明確に説明し損なうことのないいくつかの詳細を見逃してはなりません。
詳細情報:ケトジェニックダイエット (ATP)、これは「純粋なエネルギー」の唯一の真のコンテナおよびディストリビューターとして定義できます。
実際、アデノシンからリン酸を分離するたびに(ATP-> ADP、酵素ATPaseの加水分解による)、エネルギーが大幅に放出されます。この反応は、生物のすべての細胞プロセスに利用されます。したがって、基質と代謝経路によって、わずかに異なる方法ではありますが(もちろん)、エネルギー生産の最終的な目標は常に「ATP(ADP-> ATP)」の再充電です。
しかし、ATPはどのように再充電されますか?道は長いですが、私たちは最後から始めたので、すべてを逆にたどります。
あなたが依存します)。一方、これが起こるためには、電子伝達系または呼吸鎖が最初に完成しなければなりません。NADHとFADH2は、クレブス回路(以下で説明します)中にH +が豊富な酵素であり、いわゆるシトクロムのおかげで電子が放出されます。 NADHとFADH2がNAD +とFADに酸化された後、これらは電子を伝導して特定の酵素に放出し、残留H +イオンを膜に送り込み、プロトン勾配を生成します。これらのイオンの再突入は酵素によって制御されます。それ自体。ADPを再充電するためにその電気化学的可能性を利用するATPシンターゼ。
オキサロ酢酸サイクルは、酸素の存在下でのエネルギー生成の重要な段階であり、酸化的リン酸化に必要な要素(NAD +およびFAD-> NADHおよびFADH2)の合成に加えて、細胞の他の基本的なプロセスにも関与します。
注:これは、実際には開始と終了がないため、「サイクル」と呼ばれますが、永続的に継続する必要があります。
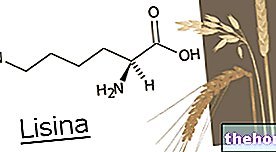
クレブス回路の主な基質はアセチルCoA(アシル基+補酵素A)であり、これは嫌気性解糖(グルコース異化作用)と脂肪酸のベータ酸化に由来します。これに細心の注意を払うことをお勧めします。それは扱われる主題の理解に不可欠だからです。
アセチルCoAのクレブス回路への侵入は、オキサロ酢酸との縮合によって起こり、クエン酸を生成します。
注意! オキサロ酢酸は、グルコースのみから生成できる分子です。その欠如は、アセチルCoAのクエン酸への縮合を危険にさらし、したがってクレブス回路に入り、アセチルCoAの蓄積を決定します。 2つのアセチルCoAの結合により、ケトン体が生成されます。
サイクル自体の終わりに、アセチルCoAによって放出された2つの炭素原子が酸化されて2つのCO2分子になり、アセチルCoAと凝縮できるオキサロ酢酸が再び再生されます。
エネルギーの観点から言えば、グアノシン三リン酸(GTP)の分子(ADPをATPに即座に再充電するために使用される)の生成です。NADHの3つの分子とFADH2の1つ(最初はNAD +とFAD)です。上で見たように、これらは、ATPシンターゼの機能を可能にするシトクロムへの酸化と移動まで電子の輸送として機能します。
次に、アセチルCoAの製造に取り掛かります。
、嫌気性糖分解の中間体。ミトコンドリアでは、ピルビン酸デヒドロゲナーゼの多酵素複合体のおかげで、これはアセチルCoAに変換されます。アセチルCoAの合成は、脂肪酸からも発生する可能性があります。これらは、細胞質で活性化され(補酵素Aの分子と結合してアシルCoA複合体を形成することにより)、「L-カルニチンの作用」のおかげでミトコンドリアマトリックスに入ります。したがって、最終的なベータ酸化が始まります。結果l "アセチル-CoA。
タンパク質アミノ酸(AA)は、アセチルCoAの生成にも使用できます。アセチルCoAは、ケトン体生成AAの脱アミノ化によって直接得られますが、クレブス回路の中間体は、糖原性AAから得られます。
肝臓と筋肉で。その欠如は、少なくとも部分的には、栄養不足の重症度と身体活動のレベルに依存しますが、グリセロール、乳酸、糖原性アミノ酸を使用してグルコースを取得する肝プロセスである新糖原性によって補われます。このため、総炭水化物の摂取量が少ないにもかかわらず、多くの高タンパク質食はケトーシスの状態を確立することを許可しません(不十分なレベルのオキサロ酢酸のみに到達することができます)。 ただし、窒素残留物は非常に高く、これにより肝臓と腎臓の作業負荷が増加します。健康な被験者では、これが適切な病状を引き起こすことはほとんどありませんが、これらの食事を長期間維持することはお勧めできません。オキサロ酢酸の不在は、細胞がケトン体の合成で改善するアセチルCoAの蓄積を決定します。幸いなことに、ケトン体はエネルギー目的にも使用でき、健康な有機体の過剰分は、尿中排泄、汗、肺の換気によって補われます。これは、特にかなりの運動活動が存在する場合に、体が全能力で機能することを意味するものではありません。
さらに、これらが食欲の部分的な抑制を間違いなく決定したとしても(ケトン食療法に関する記事を参照)、この利点は、脂肪酸の細胞使用を減らすという副作用によって無効になります。
一方、1型糖尿病、腎不全、肝不全などの病気の被験者では、重度の病的ケトアシドーシスが発症する可能性があります。
詳細:減量のための生理的ケトーシス:それらは機能しますか?