有糸分裂は通常、前期、中期、後期、終期と呼ばれる4つの期間に分けられます。その後、細胞形成と呼ばれる2つの娘細胞に分裂します。
前期
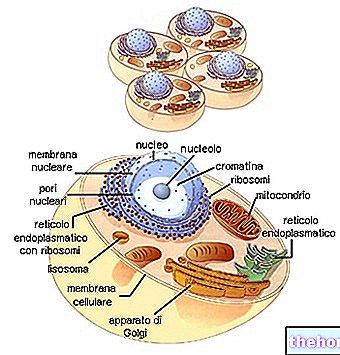
核の中には、まだ細長く、ボールに巻かれた着色可能なフィラメントの描写が徐々に見られます。したがって、核タンパク質に結合したDNA鎖が徐々にらせん状になると、染色体が徐々に識別可能になります。その間、核小体は消え、中心小体は2倍になります。核膜の溶解が始まる間、2つの中心小体は核の反対の極に移動します。前期から中期への移行の瞬間(一部は前中期として別々に識別されるため)、染色体は短くなり、はっきりと見え、核膜で分離されなくなります。中心小体は反対の極にあり、微小管の紡錘体がそれらを子午線様式に接続しています。核小体は溶解しています。中心小体をつなぐ紡錘体は、(染色体とは異なり)着色できないため無彩色紡錘体と呼ばれ、連続繊維(紡錘繊維)と、中点で染色体の中心小体につながる繊維(染色体繊維)の両方が含まれます。

中期
中期では紡錘体が明確に区別され、すべての染色体が赤道プレートと呼ばれる赤道面に配置されます。この瞬間、染色体は最大に短縮されます。これは、細胞が染色体を数え、識別するために固定される瞬間です。各染色体は、セントロメア(セントロメア( 2つの染色分体は複製の結果です)。セントロメアは、2つの血小板間の一種の接着のように、唯一の接触点です。すべてのセントロメアは、無彩色の紡錘体の染色体線維の中心点に付着しています(これが染色体が赤道位置にある理由です)。
後期
中期の終わりに、各セントロメアが2倍になり、各半分が紡錘体に沿ってそれぞれの極の方向に移動することに注意してください。この時点で、それぞれのセントロメアによって引きずられる染色分体は、明らかに2つのグループに分離します。このように分離された各染色分体は、いわば成人します。これからは、それぞれの娘細胞に向けられた染色体になります。
終期
2つの同一のクラスターに分離された染色体は、脱スパイラル化に戻り、2つの新しい細胞の核を再構成します。無彩色の溶融物が溶解します。
細胞形成
細胞質はまた、徐々に絞ることによって分裂し、2つの娘細胞に体積と細胞小器官の両方のそれぞれの比率に起因します。特に、各娘細胞は中心小体を受け取っているため、少なくとも1つのミトコンドリアを受け取る必要があります(前述のように、これらは独自の遺伝的連続性を持つ構造です)。
植物界では、有糸分裂の一般的な特徴は尊重されますが、いくつかの違いがあることに注意する必要があります。まず第一に、中心小体が欠落しています。紡錘体の極には、中心体と呼ばれる光学的に空の空間があり、そこから微小管が放射します。さらに、色素体の帰属(その遺伝的連続性による)も各娘細胞に保証されなければならない細胞分化の際に、娘細胞の分離は、絞扼によってではなく、中隔の形成によって起こる。最初は原形質膜のみで、次に細胞壁が介在します。
遺伝的基盤は、「遺伝物質の倍増(DNAの複製、つまり、セントロメアを介して結合された2つの等しい染色分体への各染色体の倍増)と半分(セントロメアの分離、移動)の間の交代によって表されます。 2つの新しい等しい核を構成するための反対方向の2つの染色分体の)。
後でわかるように、染色体は同族体のペア(それぞれ配偶子から来る)で存在するため、分割直後の染色体鎖の数は、染色体のタイプごとに1ペアであることがわかります。単一種に特徴的な異なるタイプの染色体の数をnと呼ぶと、有糸分裂後の正常な染色体セットは、n対の相同染色体(2n染色体=二倍体細胞)です。
ただし、S期の後、各染色体は2倍になります。実際、各娘細胞に2n染色体を与えることができるようにするには、4n染色分体が必要です。このようにして、DNAの4n鎖と2n鎖の間で複製と有糸分裂が交互に起こることがわかります。